Andy
Senior Member (Voting rights)
@DMissa , is it this research that you will be talking about at UK: Conference: MitOX 2024, 12th April 2024
I have long been of the view that most of what we see and know so far about ME is the patient's physiological and behavioural reaction to the underlying problem, the attempts to deal with a pathological demand on otherwise normal healthy systems now being forced to operate beyond their sustainable capacity, not the underlying problem itself.I've never thought a defect in energy production was the main cause of ME/CFS. I think energy production is impaired because the cells are shifting into a "defensive" state for some reason.
@DMissa , is it this research that you will be talking about at UK: Conference: MitOX 2024, 12th April 2024
Making a hole in somebody's breast bone is not trivial so before doing a bone marrow one really needs to have some idea what one might be looking for and what it might explain. With nothing very obvious being present in circulating cells it is unclear what one might find.
Moreover, hundreds of people with ME will have had bone marrows over the years for other reasons and nobody has noticed anything unusual. Bone marrow contains different areas with different sorts of cells in varying proportions, so one cannot get much useful numerical data from it, except perhaps in terms of maturation of specific cell lines. You would need about 100 samples to begin to pick up anything statistical I think.
The problem with bone marrows done for other reasons is that the marrow may be abnormal for those other reasons, so unless something new, unique to ME, is found, it isn't much help, and there hasn't been.
It makes sense to consider marrow and Jo Cambridge and I would always consider that if trying to piece together a theory about immune cells, but even in other conditions where you have more to go on it is rarely a viable option. We never did a formal research bone marrow study in autoimmune disease.
- CD38 is known to degrade NAD+, removing the NAD+ pool would reduce the speed of ATP production, do you think Daratumumab may have potential by simply removing cells that are more likely to reduce the NAD+ pool?
- CD24 seems to have been identified consistently elevated on B cells in ME cohorts by Jo Cambirdge, no one else is yet to even look at it, what is your opinion on why that could be?
I don't really see the connection. Like CD20, CD38 is just a useful ligand for identifying cells for antibody-mediated killing. If you kill lots of plasma cells you end up with no antibody production but not much effect on the rest of the body as far as I know.
I see that CD38 seems to be a significant NADase, but I am unclear what the significance of that is to metabolic function. In 2000 we didn't use anti-CD38 because we were told CD38 was expressed on other cells, such as cardiomyocytes. It seems to be that anti-CD38 antbodies do not cause major problems with these other cells. So presumably their metabolism carries on as usual.
Jo was interested in CD24 because in repopulation following rituximab CD24 return behaved strangely. I think particularly in patients with thrombotic thrombocytopenia purpura. There aren't really any other B cell biologists working on ME/CFS as far as I know, now that Amok Bansal has largely retired from research. Even most other immunologists probably have little interest in CD24.
If B cells are relevant to ME/CFS I suspect the story is subtle and not a matter of some obscure specific autoantibody effect. Something that has proven important in lupus is the control of extra follicular B cell survival. It may also be relevant in RA in a different way. The question arises as to whether in ME/CFS the female predominance reflects a shift in maturation control for B cells, analogous to but not the same as in lupus, that allows for the formation of antibody populations that are not autoreactive but 'bad' for some other generic reason. If they were directed against foreign antigens and made by long lived plasma cells then rituximab would make no difference.
Maybe there is another 'quality control' mechanism for antibodies that is defective in people with ME/CFS such that they form antibodies to microbes that get in the way in some non-specific way - maybe by forming complexes of the wrong size or triggering too much ADCC or something. These antibodies might normally be selected against during an immune response through some interaction with CD57+ T cells outside follicles or something (just an illustration).
CD24 might be a marker of such a maturation shift, which might of course also be reflected in a shift in mean values for B cell metabolic pathways.
which may be due to the underlying problem or a reflection of that problem.
Do you think this increased expression of CD38 on stimulated B cells could have any link to the Daratumumab trial results?
Hallo @Jonathan Edwards, @DMissa,
Do you think this increased expression of CD38 on stimulated B cells could have any link to the Daratumumab trial results?
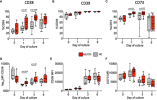
Because it looks like the % difference on CD38 is quite large. For example, in the chart below, on day 3 of culture of the B cells, whatever that means, the median HC CD38 level was 45% but the median ME CD38 level was 75%. That's a large difference.
To me this links to two things:
1. Dara is potentially depleting some B cells since they have CD38 in ME patients. But this is not important as Rituximab already depletes B cells without much effect.
2. Somehow the ME/CFS LLPCs have more CD38. Which makes them better targets for Dara. I think this is a very hopeful stretch. Any LLPCs that came from B cells, post-ME, "inherit" this higher expression of CD38.
Could this be a reasonable pulling together of different findings:
I think that would be a fair suggestion. It needn't involve any inflammation, just antibodies misbehaving in signalling systems. It would fit with the FcR1 idea, although I don't have any great investment in that other than as an example.
It would be nice if TLR7 or 8 could be involved, although BTK being on the X chromosome might be enough to explain increased risk in women.
One thing Jo and I talked about in relation to unwinding autoimmune loops was that it would be good to discourage ongoing re-education of new B cell clones by old B cells clones through blocking a signalling route rather than just having to kill everything as for ritux. If Dara were to dispose of long lived plasma cells producing unwanted antibody AND block ongoing re-education of B clones destined to make new unwanted antibody the 'reset' button might do its job.
NK cells might be important for plasma cell killing but not needed for the other bit.
That of course was in the context of traditional autoimmunity but we always assumed that there would be variants on the theme that did not necessarily involve classical autoantibodies - maybe including MS.
Are there any drugs that raise NK cells that could be used prior to Dara? Like a combo treatment.
Can you define what you mean by inflammation is this context? Is seems to mean something different to every person, so it’s difficult to keep track.I'm not saying that ME/CFS has ongoing inflammation, I'm more suggesting that PVFS does and it might be that what makes ME/CFS is priming the B cells to have more sensitive CD38, which in turn increases the pathogenic plasma cells.
Now the same thing to prime the CD38 also opens up more niches for plasma cells. This chronic inflammation could be set up by having a chronic infection phase or a drawn out acute phase. Perhaps it starts with post viral fatigue syndrome but then those that get ME/CFS are the ones that get this CD38 priming and in appropriate plasma cells in more niches. Those that don't get the priming and niches simply recover from the PVFS and never meet ME/CFS.
What is hard for me to reconcile about this proposed higher CD38 induction —> more plasma cells problem is that it would only make sense if you also had a magic mechanism like in the Fcgr1 hypothesis explaining how the antibodies themselves, once produced by these plasma cells, invoke a response that leads to ME/CFS symptoms without all the simultaneous inflammation-inducing signaling pathways that you normally get from antibody binding and immune complex stimulation.I think that would be a fair suggestion. It needn't involve any inflammation, just antibodies misbehaving in signalling systems. It would fit with the FcR1 idea, although I don't have any great investment in that other than as an example.
explaining how the antibodies themselves, once produced by these plasma cells, invoke a response that leads to ME/CFS symptoms without all the simultaneous inflammation-inducing signaling pathways that you normally get from antibody binding and immune complex stimulation.
Sure but in those cases we see progressive tissue damage and [edit: constant] differences in immune populations even just doing a single cell screen of circulating or local cells at the site of damage. My point is just that we have no apparent sign whatsoever of any abnormality that can be traced back to antibodies other than the feeling of being sick. [Edit: it would be a magic bullet solution indeed if there was a way that antibodies caused a problem that only lead to the feeling of being sick, by a completely different and unknown mechanism than the ones we already know about that mediate sickness behavior during infection.]Worth remembering that in many autoimmune diseases antibodies do not cause inflammation. They do other things. RA is the paradigmatic small immune complex disease, involving TNF, but lupus is mediated through a whole lot of other pathways, including effects of large complexes that do not appear to involve TNF (CRP is often normal). And in conditions like scleroderma the problem is unlikely to be immune complexes at all. Antibodies may be interacting with topoisomerase-1 actually within nuclei.